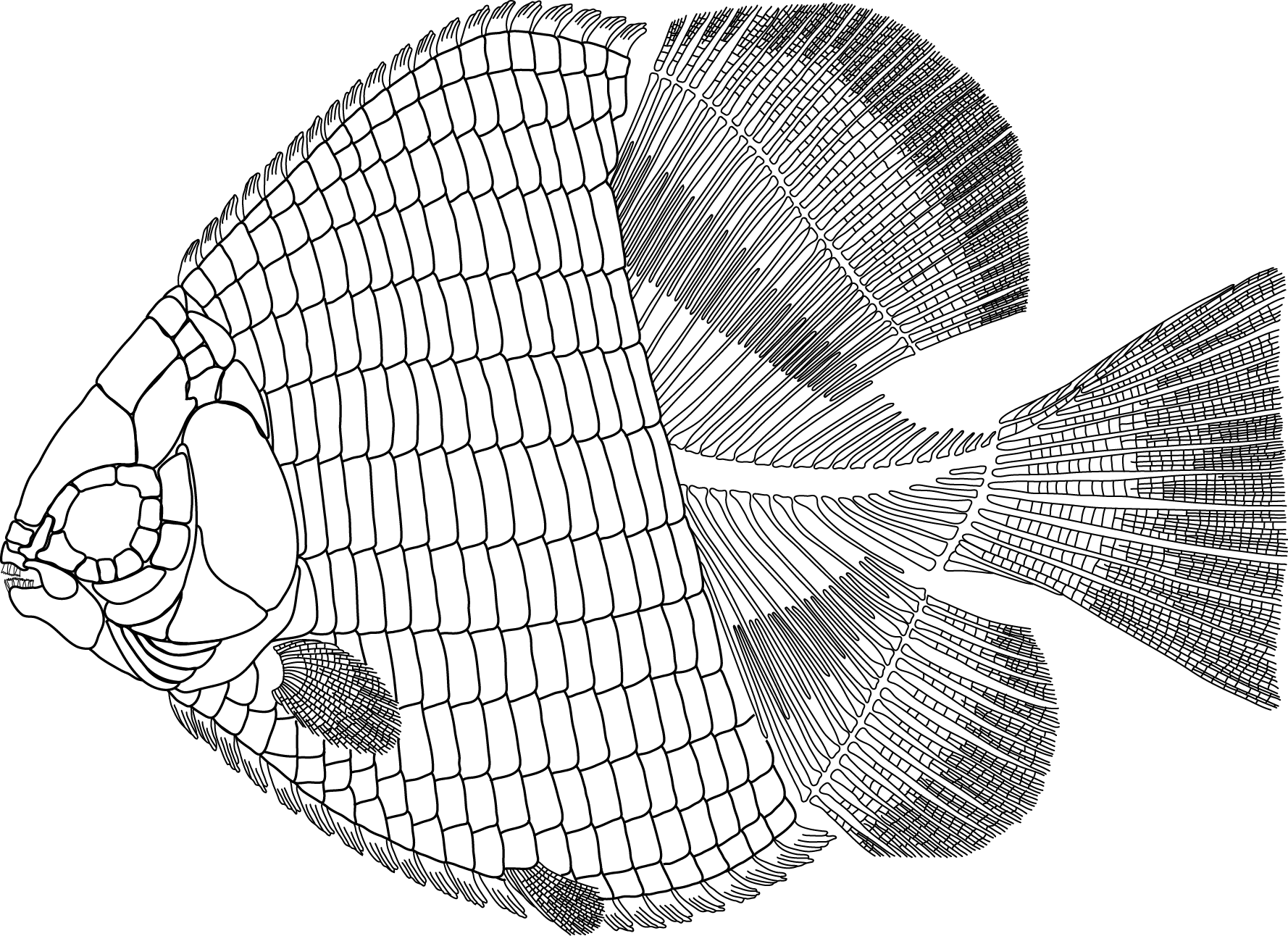

Could this fossil push the age of bony fishes further back?
Alamosaurus: how this massive titan’s neck is impacting relationships of titanosaurs | PLOS Paleo Community

This is a blog post I originally wrote for the PLOS Paleontology Community blog, and am archiving it here on my personal website. You can find the original post here. Remember the Alamo? Well, it’s easy to forget when you are staring at this massive dinosaur. It makes that Tyrannosaurus look like a puppy in comparison.Continue reading "Alamosaurus: how this massive titan’s neck is impacting relationships of titanosaurs | PLOS Paleo Community"
Track Makers in Southern Utah: The St. George Dinosaur Discovery Site | PLOS Paleo Community

Southern Utah boasts one of the best dinosaur tracksites in North America. Here's why you should visit it.
Paleo in PLOS: November 2019
Hey folks! Sarah here. Since this weekend is a holiday in the United States, I'm throwing together this latest list of paleontological research quickly. I hope you all are recovering from your Thanksgiving dinosaur carcass consumption coma and getting ready to embrace a chilly winter (at least in the northern hemisphere)! Here is a listContinue reading "Paleo in PLOS: November 2019"
It’s a Hard-Knock Life for an Ichthyosaur | PLOS Paleo Community

A new study published last week in PLOS ONE highlights just how many broken bones and traumatic injuries and infections a species of ichthyosaur can take.
Munching and Migrating Megabeasts: Sauropod teeth illuminate migration patterns between Europe and Africa

What do sauropod teeth tell us about faunal connections in Europe and Africa?
Pregnant Plesiosaurs and Baby Bones: Bone histology reveals ontogeny in polycotylid plesiosaurs — PLOS Paleo Community
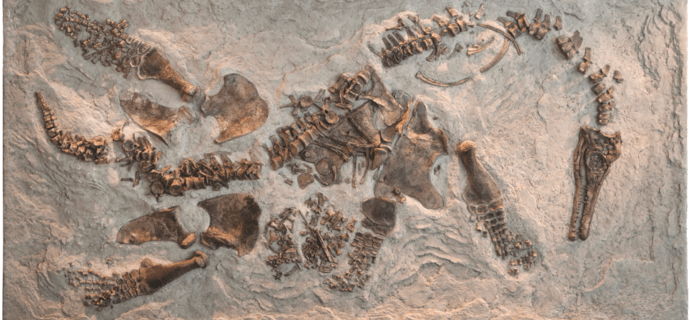
Pregnancy in the fossil record is an exciting find. Setting aside the sad fact that an unfortunate mother met her demise while carrying a baby, these one in a million specimens provides some key insight into the behavior and lifestyle of organisms unlike any living today. One such specimen is on display at the Natural HistoryContinue reading "Pregnant Plesiosaurs and Baby Bones: Bone histology reveals ontogeny in polycotylid plesiosaurs — PLOS Paleo Community"
Paleo in PLOS: October 2019
Featured image: reconstruction of a Late Cretaceous dinosaur tracksite in Alaska. From Fiorillo et al. (2019), see below Happy Halloween! PLOS has seen a lot of activity in the paleo realm this month, including a lot of exciting discoveries! So let's dive right into the latest published research across all PLOS journals.... Competition-driven evolution ofContinue reading "Paleo in PLOS: October 2019"
Paleo in PLOS: September 2019
PLOS saw some great paleontological papers published this month. Let's highlight some of the work that was published in the freely accessible PLOS ONE in September. For a complete and regularly updated list of open access papers being published in dozens of journals, be sure to follow our regularly updated Twitter feed. Resource partitioning among brachiopodsContinue reading "Paleo in PLOS: September 2019"
About the Fossil Friday Roundup…
Hey folks! Sarah here. I just wanted to send a quick message for those of you looking for the Fossil Friday Roundup. The roundup takes a fair amount of time to compile each week, and I have been finding it increasingly harder to keep up with posting it in recent weeks. If you enjoy catchingContinue reading "About the Fossil Friday Roundup…"
