Featured Image: Ngwevu intloko: a new early sauropodomorph dinosaur from the Lower Jurassic Elliot Formation of South Africa. From Chapelle et al. (2019). Papers (All Open Access): Synchronizing volcanic, sedimentary, and ice core records of Earth’s last magnetic polarity reversal (Science Advances) Pliocene warmth consistent with greenhouse gas forcing (Geophysical Research Letters) Early animal evolution: aContinue reading "Fossil Friday Roundup: August 9, 2019"
Fossil Friday Roundup: July 26, 2019
Featured Image: Seazzadactylus venieri, from Dalla Vecchia (2019). Papers (All Open Access): Aloisalthella, a new genus of fossil Polyphysacean green algae (Chlorophyta, Dasycladales), with notes on the genus Clypeina (Michelin, 1845) (PalaeoE) An annotated catalogue of types of Silurian–Devonian brachiopod species from southern Belgium and northern France in the Royal Belgian Institute of Natural Sciences (1870–1945), withContinue reading "Fossil Friday Roundup: July 26, 2019"
Fossil Friday Roundup: July 19, 2019
Featured Image: Odontochelys semitestacea, from Schoch et al. (2019). Papers (All Open Access): Crustose lichens with lichenicolous fungi from Paleogene amber (SciRep) Ricciopsis sandaolingensis sp. nov., a new fossil bryophyte from the Middle Jurassic Xishanyao Formation in the Turpan-Hami Basin, Xinjiang, Northwest China (PalaeoE) Frond morphology and epidermal anatomy of Compsopteris wongii (T. Halle) Zalessky fromContinue reading "Fossil Friday Roundup: July 19, 2019"
Fossil Friday Roundup: July 12, 2019
Featured Image: From Hartman et al. (2019). Papers (All Open Access): Revisiting metazoan phylogeny with genomic sampling of all phyla (Link) First cycad seedling foliage from the fossil record and inferences for the Cenozoic evolution of cycads (Biology Letters) An Evolutionarily Ancient Immune System Governs the Interactions between Pseudomonas syringae and an Early-Diverging Land PlantContinue reading "Fossil Friday Roundup: July 12, 2019"
Fossil Friday* Roundup: July 6, 2019
*Saturday edition Featured Image: Right lateral view of the Cioclovina calvaria exhibiting a large depressed fracture. From Kranioti et al. (2019). Papers (All Open Access): Accelerated diversifications in three diverse families of morphologically complex lichen-forming fungi link to major historical events (SciRep) Preliminary investigation of a diverse megafossil floral assemblage from the middle Miocene of southernContinue reading "Fossil Friday* Roundup: July 6, 2019"
Fossil Friday Roundup: June 28, 2019
Featured Image: From Langer et al. (2019). Papers (All Open Access): Nonequilibrium evolution of volatility in origination and extinction explains fat-tailed fluctuations in Phanerozoic biodiversity (Science Advances) Biodiversity, Disparity and Evolvability (Link) Mediterranean Geoscience Reviews: a Mediterranean perspective to geosciences (MGR) Experimental evidence for species-dependent responses in leaf shape to temperature: Implications for paleoclimate inferenceContinue reading "Fossil Friday Roundup: June 28, 2019"
Fossil Friday Roundup: June 21, 2019
Featured Image: Isisfordia molnari sp. nov., a new basal eusuchian from the mid-Cretaceous of Lightning Ridge, Australia From Hart et al. 2019. Papers (All Open Access): Microbially induced potassium enrichment in Paleoproterozoic shales and implications for reverse weathering on early Earth (Nature Communications) A tectonically driven Ediacaran oxygenation event (Nature Communications) A Continuous Palynological Record ofContinue reading "Fossil Friday Roundup: June 21, 2019"
Fossil Friday Roundup: June 14, 2019
Featured Image: From Hausmann et al. (2019). Papers (All Open Access): Lunar cycles and rainy seasons drive growth and reproduction in nummulitid foraminifera, important producers of carbonate buildups (SciRep) Quantifying spatial variability in shell midden formation in the Farasan Islands, Saudi Arabia (PLOS ONE) Trilobite compound eyes with crystalline cones and rhabdoms show mandibulate affinitiesContinue reading "Fossil Friday Roundup: June 14, 2019"
Fossil Friday Roundup: June 7, 2019
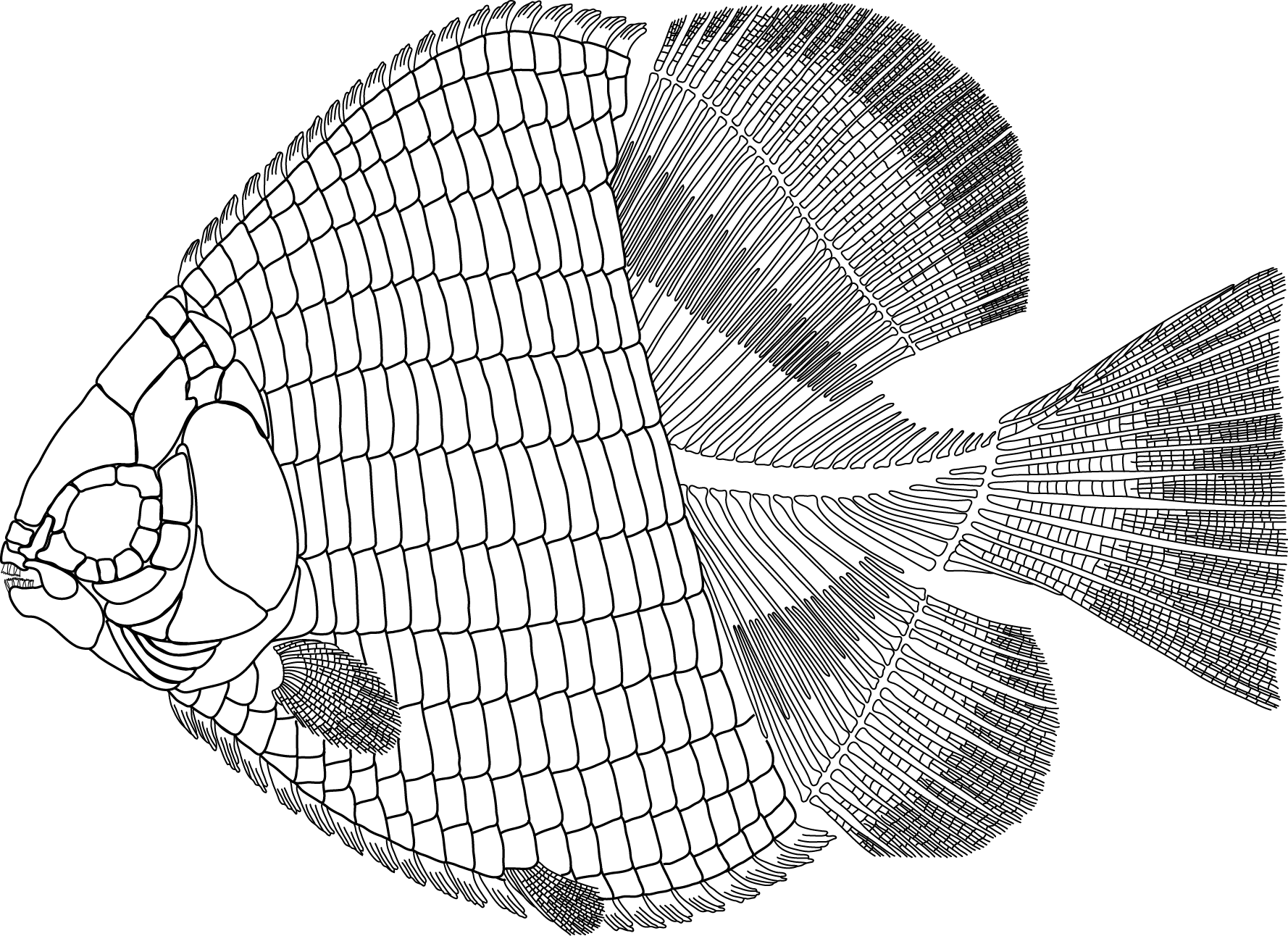
Featured Image: Coreoperca maruoi Yabumoto and Uyeno, 2009 KMNH VP 100,261. From Yabomuto (2019). Papers (All Open Access): Acta Palaeontologica Polonica 64 (2) 2019: Complete Open Access Journal Issue Timing and ecological priority shaped the diversification of sedges in the Himalayas (PeerJ) An ammonite trapped in Burmese amber (PNAS) An ichthyodectiform fish, Amakusaichthys goshouraensis (newContinue reading "Fossil Friday Roundup: June 7, 2019"
Fossil Friday Roundup: May 31, 2019
Featured Image: Fossilized school of fish. From Mizumoto et al. (2019). Papers (All Open Access): The Effect of Fossil Sampling on the Estimation of Divergence Times With The Fossilised Birth Death Process (Systematic Biology) Swapping Birth and Death: Symmetries and Transformations in Phylodynamic Models (Systematic Biology) An overview of Open Access publishing in palaeontology (PalaeoE)Continue reading "Fossil Friday Roundup: May 31, 2019"
